
Page 57 - Read Online
P. 57
Page 352 vonderEmbse et al. Neuroimmunol Neuroinflammation 2020;7:345-59 I http://dx.doi.org/10.20517/2347-8659.2019.29
A B
C D
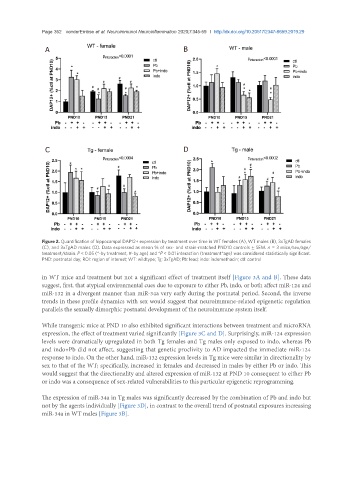
Figure 2. Quantification of hippocampal DAP12+ expression by treatment over time in WT females (A), WT males (B), 3xTgAD females
(C), and 3xTgAD males (D). Data expressed as mean % of sex- and strain-matched PND10 controls ± SEM. n = 3 mice/sex/age/
treatment/strain. P < 0.05 (*-by treatment, #-by age) and *P < 0.01 interaction (treatment*age) was considered statistically significant.
PND: postnatal day; ROI: region of interest; WT: wildtype; Tg: 3xTgAD; Pb: lead; indo: indomethacin; ctl: control
in WT mice and treatment but not a significant effect of treatment itself [Figure 3A and B]. These data
suggest, first, that atypical environmental cues due to exposure to either Pb, indo, or both affect miR-124 and
miR-132 in a divergent manner than miR-34a very early during the postnatal period. Second, the inverse
trends in these profile dynamics with sex would suggest that neuroimmune-related epigenetic regulation
parallels the sexually dimorphic postnatal development of the neuroimmune system itself.
While transgenic mice at PND 10 also exhibited significant interactions between treatment and microRNA
expression, the effect of treatment varied significantly [Figure 3C and D]. Surprisingly, miR-124 expression
levels were dramatically upregulated in both Tg females and Tg males only exposed to indo, whereas Pb
and indo+Pb did not affect, suggesting that genetic proclivity to AD impacted the immediate miR-124
response to indo. On the other hand, miR-132 expression levels in Tg mice were similar in directionality by
sex to that of the WT; specifically, increased in females and decreased in males by either Pb or indo. This
would suggest that the directionality and altered expression of miR-132 at PND 10 consequent to either Pb
or indo was a consequence of sex-related vulnerabilities to this particular epigenetic reprogramming.
The expression of miR-34a in Tg males was significantly decreased by the combination of Pb and indo but
not by the agents individually [Figure 3D], in contrast to the overall trend of postnatal exposures increasing
miR-34a in WT males [Figure 3B].