
Page 657 - Read Online
P. 657
Tafur et al. J Cancer Metastasis Treat 2018;5:xx I http://dx.doi.org/10.20517/2394-4722.2018.102 Page 9 of 14
A B
Ki67 Cyclin B1
C Cleaved PARP PCNA STK15
Cleaved PARP
β-actin
D E
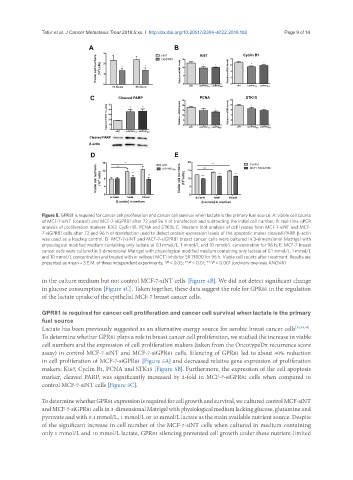
Figure 5. GPR81 is required for cancer cell proliferation and cancer cell survival when lactate is the primary fuel source. A: viable cell counts
of MCF-7-siNT (control) and MCF-7-siGPR81 after 72 and 96 h of transfection and subtracting the initial cell number; B: real-time qPCR
analysis of proliferation markers: Ki67, Cyclin B1, PCNA and STK15; C: Western blot analysis of cell lysates from MCF-7-siNT and MCF-
7-siGPR81 cells after 72 and 96 h of transfection used to detect protein expression levels of the apoptotic maker cleaved-PARP. β-actin
was used as a loading control; D: MCF-7-siNT and MCF-7-siGPR81 breast cancer cells were cultured in 3-dimensional Matrigel with
physiological modified medium containing only lactate at 0.1 mmol/L, 1 mmol/L and 10 mmol/L concentration for 96 h; E: MCF-7 breast
cancer cells were cultured in 3-dimensional Matrigel with physiological modified medium containing only lactate at 0.1 mmol/L, 1 mmol/L
and 10 mmol/L concentration and treated with or without MCT1 inhibitor SR 13800 for 96 h. Viable cell counts after treatment. Results are
presented as mean ± S.E.M. of three independent experiments; *P < 0.05; **P < 0.01; ***P < 0.001 (ordinary one-way ANOVA)
in the culture medium but not control MCF-7-siNT cells [Figure 4B]. We did not detect significant change
in glucose consumption [Figure 4C]. Taken together, these data suggest the role for GPR81 in the regulation
of the lactate uptake of the epithelial MCF-7 breast cancer cells.
GPR81 is required for cancer cell proliferation and cancer cell survival when lactate is the primary
fuel source
Lactate has been previously suggested as an alternative energy source for aerobic breast cancer cells [13,14,34] .
To determine whether GPR81 plays a role in breast cancer cell proliferation, we studied the increase in viable
cell numbers and the expression of cell proliferation makers (taken from the OncotypeDx recurrence score
assay) in control MCF-7-siNT and MCF-7-siGPR81 cells. Silencing of GPR81 led to about 40% reduction
in cell proliferation of MCF-7-siGPR81 [Figure 5A] and decreased relative gene expression of proliferation
makers; Ki67, Cyclin B1, PCNA and STK15 [Figure 5B]. Furthermore, the expression of the cell apoptosis
marker, cleaved PARP, was significantly increased by 2-fold in MCF-7-siGPR81 cells when compared to
control MCF-7-siNT cells [Figure 5C].
To determine whether GPR81 expression is required for cell growth and survival, we cultured control MCF-siNT
and MCF-7-siGPR81 cells in 3-dimensional Matrigel with physiological medium lacking glucose, glutamine and
pyruvate and with 0.1 mmol/L, 1 mmol/L or 10 mmol/L lactate as the main available nutrient source. Despite
of the significant increase in cell number of the MCF-7-siNT cells when cultured in medium containing
only 1 mmol/L and 10 mmol/L lactate, GPR81 silencing prevented cell growth under these nutrient-limited