
Page 34 - Read Online
P. 34
Garro-Núñez et al. J Transl Genet Genom 2022;6:361-74 https://dx.doi.org/10.20517/jtgg.2022.10 Page 363
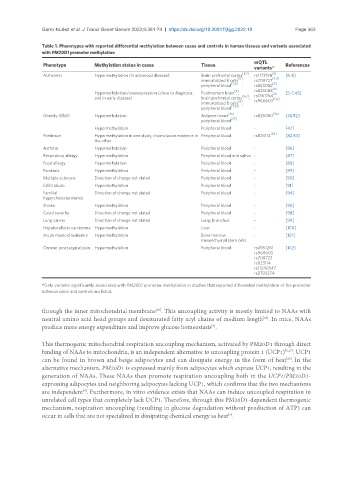
Table 1. Phenotypes with reported differential methylation between cases and controls in human tissues and variants associated
with PM20D1 promoter methylation
mQTL
Phenotype Methylation status in cases Tissue References
variants*
[6,7] [5]
Alzheimer Hypermethylation (in advanced disease) Brain prefrontal cortex , rs1172198 [6-8]
[6]
immortalized B cells , rs708727 [5-8]
[7,8] [5]
peripheral blood rs823082
[5]
[5] rs823088
Hypomethylation/overexpression (close to diagnosis Postmortem brain , [5] [5-7,45]
[6,7] rs1361754
and in early disease) brain prefrontal cortex , [5,6]
[6] rs960603
immortalized B cells ,
peripheral blood [7,45]
[36] [36]
Obesity (BMI) Hypomethylation Adipose tissue , rs823080 [36,52]
peripheral blood [52]
Hypermethylation Peripheral blood - [47]
Parkinson Hypermethylation in one study, inconclusive evidence in Peripheral blood rs823114 [83] [82,83]
the other
Asthma Hypomethylation Peripheral blood - [86]
Respiratory allergy Hypermethylation Peripheral blood and saliva - [87]
Food allergy Hypomethylation Peripheral blood - [88]
Psoriasis Hypermethylation Peripheral blood - [89]
Multiple sclerosis Direction of change not stated Peripheral blood - [90]
Child abuse Hypermethylation Peripheral blood - [91]
Familial Direction of change not stated Peripheral blood - [94]
hypercholesterolemia
Stroke Hypermethylation Peripheral blood - [96]
Covid severity Direction of change not stated Peripheral blood - [98]
Lung cancer Direction of change not stated Lung, bronchus - [99]
Hepatocellular carcinoma Hypermethylation Liver - [100]
Acute myeloid leukemia Hypermethylation Bone marrow - [101]
mesenchymal stem cells
Chronic postsurgical pain Hypermethylation Peripheral blood rs4951261 [102]
rs960603
rs708723
rs823114
rs11240547
rs2793374
*Only variants significantly associated with PM20D1 promoter methylation in studies that reported differential methylation of the promoter
between cases and controls are listed.
through the inner mitochondrial membrane . This uncoupling activity is mostly limited to NAAs with
[27]
[23]
neutral amino acid head groups and desaturated fatty acyl chains of medium length . In mice, NAAs
produce more energy expenditure and improve glucose homeostasis .
[9]
This thermogenic mitochondrial respiration uncoupling mechanism, activated by PM20D1 through direct
binding of NAAs to mitochondria, is an independent alternative to uncoupling protein 1 (UCP1) [9,25] . UCP1
can be found in brown and beige adipocytes and can dissipate energy in the form of heat . In the
[28]
alternative mechanism, PM20D1 is expressed mainly from adipocytes which express UCP1, resulting in the
generation of NAAs. These NAAs then promote respiration uncoupling both in the UCP1/PM20D1-
expressing adipocytes and neighboring adipocytes lacking UCP1, which confirms that the two mechanisms
[9]
are independent . Furthermore, in vitro evidence exists that NAAs can induce uncoupled respiration in
unrelated cell types that completely lack UCP1. Therefore, through this PM20D1-dependent thermogenic
mechanism, respiration uncoupling (resulting in glucose degradation without production of ATP) can
occur in cells that are not specialized in dissipating chemical energy as heat .
[9]